
Coleoidea
| Coleoidea Temporal range: Devonian or
| |
|---|---|

| |
| A cuttlefish | |

| |
| A common octopus | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Mollusca |
| Class: | Cephalopoda |
| Clade: | Neocephalopoda |
| Subclass: | Coleoidea Bather, 1888 |
| Divisions | |
| |
Coleoidea[1][2] or Dibranchiata is one of the two subclasses of cephalopods containing all the various taxa popularly thought of as "soft-bodied" or "shell-less" (i.e. octopus, squid and cuttlefish). Unlike its extant sister group Nautiloidea, whose members have a rigid outer shell for protection, the coleoids have at most an internal shell called cuttlebone or gladius that is used for buoyancy or as muscle anchorage. Some species, notably incirrate octopuses, have lost their internal shell altogether, while in some it has been replaced by a chitinous support structure.
The major divisions of Coleoidea are based upon the number of arms or tentacles and their structure. The extinct and most primitive form, the Belemnoidea, presumably had ten equally-sized arms in five pairs numbered dorsal to ventral as I, II, III, IV and V. More modern species either modified or lost a pair of arms. The superorder Decapodiformes has arm pair IV modified into long tentacles with suckers generally only on the club-shaped distal end. Superorder Octopodiformes has modifications to arm pair II; it is significantly reduced and used only as a sensory filament in the Vampyromorphida, while Octopoda species have totally lost that arm pair. The inner surface of the suckers (acetabulum) are reinforced with rigid sucker rings which are smooth in Sepiolida, have blunt teeth in Sepiida and sharply pointed teeth in Loliginidae and Oegopsida. The arms and/or tentacles of some oegopsid families have also evolved claw-like hooks.[3]
Evolutionary history
[edit]The earliest certain coleoids are known from the Mississippian sub-period of the Carboniferous Period, about 330 million years ago. Some older fossils have been described from the Devonian,[4] but paleontologists disagree about whether they are coleoids.[5] Other cephalopods with internal shells, which could represent coleoids but may also denote the independent internalization of the shell, are known from the Silurian.[6] It has been hypothesized that the Early–Middle Cambrian fossil Nectocaris represents a coleoid (or other cephalopod) that lost its shell, possibly secondarily,[7][8] although it is more likely that Nectocaris represents an independent lineage within the Lophotrochozoa.[9]
By the Carboniferous, coleoids already had a diversity of forms, but the major radiation happened during the Tertiary.[10] Although most of these groups are traditionally classified as belemnoids, the variation among them suggests that some are not closely related to belemnites.[11]
Classification
[edit]- Class Cephalopoda
- Subclass Nautiloidea: nautilus
- Subclass †Ammonoidea: ammonites
- Subclass Coleoidea
- Division †Belemnoidea: extinct belemnoids
- Genus †Jeletzkya
- Order †Hematitida
- Order †Phragmoteuthida
- Order †Donovaniconida
- Order †Aulacocerida
- Order †Belemnitida
- Division Neocoleoidea
- Superorder Decapodiformes
- Order Bathyteuthida
- Order †Belemnitida
- Order †Diplobelida
- Order Idiosepida
- Order Myopsida – coastal squid
- Order Oegopsida – neritic squid
- Order Sepiida – cuttlefish, pygmy, bobtail and bottletail squid
- Order Spirulida – ram's horn squid
- Superorder Octopodiformes
- Family †Trachyteuthididae (incertae sedis)
- Order Vampyromorphida: vampire squid
- Order Octopoda: octopus
- Superorder Palaeoteuthomorpha
- Order †Boletzkyida
- Superorder Decapodiformes
- (uncertain order)
- family †Ostenoteuthidae[12]
- Division †Belemnoidea: extinct belemnoids
Reproduction
[edit]The majority of coleoid species are semelparous; dying after reproducing once, with males dying after insemination, and females dying after laying/brooding their clutch.[13] A few species do not conform to this trend however; vampire squid, large Pacific striped octopus, and the West Atlantic scaled squid are thought to be iteroparous.[14][15][16] This is somewhat comparable to extant nautilus, which are also iteroparous, being long-lived animals.[13][17]
Some authors prefer to separate coleoid reproduction using other terms:[13]
1. One-time reproduction (formerly semelparity); being the occurrence of "simultaneous terminal spawning" (terminal: occuring at the end of life). This group is characterized by synchronous ovulation (the ova all ripen prior to spawning), single-cycle spawning, and the absence of growth between egg batches.
2. Multiple reproductive events (formerly iteroparity). This category is divided further into:
(i) polycyclic spawning; where single egg batches/clutches develop and are laid multiple times during the spawning season, with growth occurring between production of egg batches and breeding seasons and the gonads regenerate/ripen between clutches, e.g. Nautilus ; (ii) multiple spawning; where multiple clutches of eggs develop simultaneously, which can be differentiated in the ovaries through their development stage (also known as group-synchronous ovulation). This method is defined by monocyclic spawning and the mother's growth between egg batches, e.g. Octopus chierchiae, Sthenoteuthis oualaniensis, Ommastrephes bartramii, and Dosidicus gigas ; (iii) intermittent terminal spawning; with group-synchronous ovulation, monocyclic spawning, but the mother does not somatically grow between egg batches, e.g. Loligo vulgaris subspp., Loligo bleekeri, Loligo forbesii, Illex coindetii, Todaropsis eblanae, Todarodes angolensis, and most populations of Sepia officinalis ; (iv) continuous spawning; where egg cells developed without any apparent "batches", with all stages of development potentially being present (or asynchronous ovulation), monocyclic spawning and growth between egg batches, e.g. Cirrothauma murrayi, Opisthoteuthis agassizii, Opisthoteuthis grimaldii and Grimpoteuthis glacialis, likely also includes Argonauta bottgeri, Argonauta hians, and Idiosepius pygmaeus.
Paralarva
[edit]Paralarvae (sg.: paralarva) are young cephalopods immediately after hatching, prior to the development of adult diagnostic features and before exhibiting a similar ecology to older members of the same species. The term was introduced by Richard E. Young and Robert F. Harman in 1988; the term "larva" had been used previously, but it fell out of use as the term implied a metamorphosis occurred where larval body parts are lost completely and adult body parts developed from some "rudiments" left in an embryonic state; in contrast, young cephalopods do not undergo metamorphosis, they mostly grow morphometrically, though as this process still transforms the animal significantly, being comparable to the development of fish young (fish larva), the term "paralarva" (para: near, almost; near-larva/almost larva) was thus coined.[18][19][20]
Paralarvae have been observed only in members of the Octopoda and Teuthida (which constitutes the modern definition of Coleoidea).[18][21] In the "iteroparous" species, the hatching of the paralarvae often heralds the death of the brooding mother.[13] Paralarvae may be planktonic, or they may remain on the bottom (demersal zone). Planktonic paralarvae remain so for a time, feeding on small food items (such as detritus) until they start their transition into their adult habitat and niche;[22] a young coleoid is termed a subadult when it displays the features diagnostic for species identification in the adult, without having to display size- or sex-specific features. An adult is thus an animal showing the diagnostic traits of its species, along with signs of sexual maturity.[18][23]
- Examples of paralarval cephalopods
-

-
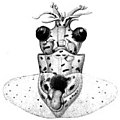
-
 Octopus paralarva
Octopus paralarva -




.jpg)

References
[edit]- ^ From Greek koleos, sheath
- ^ Nixon, Marion; Young, J. Z. (2003). The Brains and Lives of Cephalopods. New York: Oxford University Press. ISBN 978-0-19-852761-9.
- ^ Fuchs, D.; Hoffmann, R.; Klug, C. (2021). "Evolutionary development of the cephalopod arm armature: A review". Swiss Journal of Palaeontology. 140 (1): 27. Bibcode:2021SwJP..140...27F. doi:10.1186/s13358-021-00241-z. PMC 8688392. PMID 34956072.
- ^ Bandel, Klaus; Reitner, J.; Sturmer, W. (1983). "Coleoids from the Lower Devonian Black Slate ("Hunsruck-Schiefer")". Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen. 165 (3): 397–417. doi:10.1127/njgpa/165/1983/397.
- ^ Zuschin, Martin (2008). "Phylogeny and Evolution of the Mollusca". Marine Ecology. 30 (2): 269. Bibcode:2009MarEc..30..269Z. doi:10.1111/j.1439-0485.2009.00287.x.
- ^ Turek, V.; Manda, Š. (2012). "An endocochleate experiment in the Silurian straight-shelled cephalopod Sphooceras". Bulletin of Geosciences. 87 (4): 767–813. doi:10.3140/bull.geosci.1335.
- ^ Smith, M. R.; Caron, J. B. (2010). "Primitive soft-bodied cephalopods from the Cambrian". Nature. 465 (7297): 469–472. Bibcode:2010Natur.465..469S. doi:10.1038/nature09068. hdl:1807/32368. PMID 20505727. S2CID 4421029.
- ^ Smith, M. R. (2013). "Nectocaridid ecology, diversity and affinity: early origin of a cephalopod-like body plan". Paleobiology. 39 (2): 291–321. Bibcode:2013Pbio...39..297S. doi:10.1666/12029. S2CID 85744624.
- ^ Kröger, Björn; Vinther, Jakob; Fuchs, Dirk (August 2011). "Cephalopod origin and evolution: A congruent picture emerging from fossils, development and molecules: Extant cephalopods are younger than previously realised and were under major selection to become agile, shell-less predators". BioEssays. 33 (8): 602–613. doi:10.1002/bies.201100001. PMID 21681989. S2CID 2767810.
- ^ Boyle, Peter; Rodhouse, Paul (15 April 2008). Cephalopods: Ecology and Fisheries. John Wiley & Sons. ISBN 978-1-4051-4543-5.
- ^ Doguzhaeva, Larisa A.; Mapes, Royal H.; Mutvei, Harry (2007). "A Late Carboniferous Coleoid Cephalopod from the Mazon Creek Lagerstätte (USA) with a radula, arm hooks, mantle tissues, and ink". In Landman, Neil H.; Davis, Richard Arnold; Mapes, Royal H. (eds.). Cephalopods Present & Past: New Insights and Fresh Perspectives. Berkeley & Los Angeles, California, USA: University of California Press. pp. 121–143.
- ^ Garassino, A.; Donovan, D. T. (2000). "A New Family of Coleoids from the Lower Jurassic of Osteno, Northern Italy". Palaeontology. 43 (6): 1019. Bibcode:2000Palgy..43.1019G. doi:10.1111/1475-4983.00160.
- ^ a b c d Rocha, F.; Guerra, Á.; González, Á. F. (2001). "A review of reproductive strategies in cephalopods". Biological Reviews of the Cambridge Philosophical Society. 76 (3): 291–304. doi:10.1017/S1464793101005681. PMID 11569786. S2CID 5777682.
- ^ Hoving, Henk-Jan T.; Laptikhovsky, Vladimir V.; Robison, Bruce H. (20 April 2015). "Vampire squid reproductive strategy is unique among coleoid cephalopods" (PDF). Current Biology. 25 (8): R322 – R323. Bibcode:2015CBio...25.R322H. doi:10.1016/j.cub.2015.02.018. PMID 25898098. S2CID 668950. Retrieved 29 July 2020.
- ^ Caldwell, Roy L.; Ross, Richard; Rodaniche, Arcadio; Huffard, Christine L. (12 August 2015). "Behavior and Body Patterns of the Larger Pacific Striped Octopus". PLOS ONE. 10 (8): e0134152. Bibcode:2015PLoSO..1034152C. doi:10.1371/journal.pone.0134152. ISSN 1932-6203. PMC 4534201. PMID 26266543.
- ^ Hoving, H. J. T.; Vecchione, M. (December 2012). "Mating Behavior of a Deep-Sea Squid Revealed by in situ Videography and the Study of Archived Specimens". The Biological Bulletin, University of Chicago. 223 (3): 263–267. doi:10.1086/BBLv223n3p263. PMID 23264472. Retrieved 30 March 2025.
- ^ Saunders WB (June 1984). "Nautilus Growth and Longevity: Evidence from Marked and Recaptured Animals". Science. 224 (4652): 990–992. Bibcode:1984Sci...224..990S. doi:10.1126/science.224.4652.990. PMID 17731999. S2CID 40891271.
- ^ a b c Richard Edward Young & Robert F. Harman (1988). ""Larva", "Paralarva", and "Subadult" in Cephalopod Terminology". Malacologia. 29 (1): 201–207. Retrieved August 13, 2011.
- ^ Young, R.; Harman, R. ""Larva,""paralarva" and "subadult" in cephalopod terminology". semanticscholar.org. Retrieved 6 April 2025.
- ^ Zaragoza, N.; Quetglas, A.; Moreno, A.; Anderson, Emory D. (2015). Identification guide for cephalopod paralarvae from the Mediterranean Sea (PDF). Vol. 324. Copenhagen: ICES Cooperative Research Report. ISBN 978-87-7482-156-4. ISSN 1017-6195. Retrieved 6 April 2025.
- ^ Fernández-Gago, Raquel; Molist, Pilar; Anadón, Ramón (March 2019). "Tissues of Paralarvae and Juvenile Cephalopods". Handbook of Diseases and Pathogens in Cephalopods: 87–109. Retrieved 6 April 2025.
- ^ Fernández-Álvarez, Fernando Á; Machordom, Annie; García-Jiménez, Ricardo; Salinas-Zavala, César A; Villanueva, Roger (February 21 2018). "Predatory flying squids are detritivores during their early planktonic life". Sci Rep . 8. doi:10.1038/s41598-018-21501-y. PMID 29467371. Retrieved 6 April 2025.
{{cite journal}}: Check date values in:|date=(help) - ^ Vidal, Erica A. G.; Shea, Elizabeth K. (July 2023). "Cephalopod ontogeny and life cycle patterns". Frontiers in Marine Science. 10. doi:10.3389/fmars.2023.1162735. Retrieved 6 April 2025.
{{cite journal}}: CS1 maint: unflagged free DOI (link)